The RMgm database now contains information on the genotype and phenotype of more than 5300 different mutants. The phenotype data of all (new) mutants are linked to the individual gene pages of the malaria database of `EuPathDB Bioinformatics Resource Center for biodefense and Emerging / Re-emerging Infectious Diseases` (www.PlasmoDB.org).
06 Januari 2023
The RMgm database now contains information on the genotype and phenotype of more than 5200 different mutants.
25 July 2022
The RMgm database now contains information on the genotype and phenotype of 5156 different mutants.
.27 January 2022
The RMgm database now contains information on the genotype and phenotype of 5086 different mutants.
06 August 2021
The RMgm database now contains information on the genotype and phenotype of 5000 different mutants.
15 April 2021
The RMgm database now contains information on the genotype and phenotype of 4909 different mutants.
21 October 2020
The RMgm database now contains information on the genotype and phenotype of 4816 different mutants.
6 February 2020
The RMgm database now contains information on the genotype and phenotype of 4659 different mutants.
21 November 2019
Below you will find a brief description for several updates of the RMgm database.
Addition of data on mosquito and liver stage phenotypes of 1325 PlasmoGem mutants.
In a study, published recently in Cell, more than 1,300 barcoded P. berghei gene-deletion mutants were followed in a screen through the life cycle. 461 genes were discovered that are required for efficient parasite transmission to mosquitoes through the liver stage and back into the bloodstream of mice. In the RMgm database we have now added to 1325 mutants data on the change in relative abundance of the barcoded mutants in the 3 following life cycle transitions: from blood stages (B1) to midgut oocysts (MG), from midgut oocysts (MG) to salivary gland sporozoites (SG) and from salivary gland sporozoites (SG) to blood stages (B2). The change in relative abundance is classified as ‘non-reduced’, ‘reduced’ and ‘no power’. In addition, we have added the phenotypes throughout mosquito and liver stage development of 21 ‘single knockout mutants’ that were generated in the same study (for example targeting the genes biotin--protein ligase 1, long chain fatty acid elongation enzyme, putative, lipoyl synthase (LipA), phosphoacetylglucosamine mutase, putative, etc.).
Phenotypes of other new mutants in RMgmDB
A number of mutants (and their phenotypes) were recently added to the RMgm database, for example novel mutants lacking kinesins 8X and 8B, with roles in sporozoite formation (PMID: 31600347) or fertilisation (PMID: 31634979), mutants to examine function and expression of the six different myosins (PMID: 31283102), mutants lacking an alternative splicing factor with a role in sexual development (PMID: 31370870), mutants used to analyse the importance of folate metabolism in blood stages (PMID: 30625318).
13 February 2019
The RMgm database now contains information on the genotype and phenotype of >4525 different mutants.
01 October 2018
10 YEAR ANNIVERSARY RMgm DATABASE
The RMgm database (www.pberghei.eu) of genetically modified malaria parasites celebrates its 10 year Anniversary. The RMgm database was created and is maintained by the Leiden Malaria research Group. It first appeared on-line in October 2008. We would like to thank everyone who has contributed to the database and helped to improve it over the years.
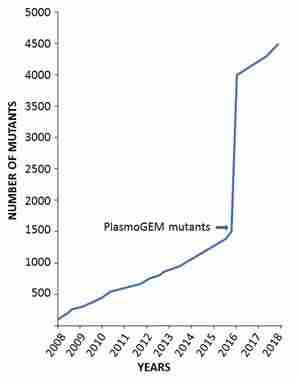
Currently, the database contains information on 4490 different genetically modified rodent malaria parasites that have been generated by many labs worldwide. It contains data on mutants of 3 rodent malaria parasites: Plasmodium berghei, P. yoelii and P. chabaudi. See below how the number of mutants published and reported in the database has rapidly increased over the past decade.
The aim of this database is to provide the research community easy access to genotype and phenotype information on published mutant malaria parasites, and thereby help to provide insights into parasite gene function and biology. In addition, it contains information on a variety of parasites expressing transgenes, for example fluorescent and luminescent proteins or transgenic parasites expressing P. falciparum or P. vivax antigens. Transgenic parasites have been used to interrogate parasite biology, host-parasite interactions and to advance malaria vaccine and drug development.
The information in this database is shared and linked to with the ‘Sanger Institute Pathogen Genome Database’ (www.GeneDB.org), ‘EuPathDB Bioinformatics Resource Center for biodefense and Emerging / Re-emerging Infectious Diseases’ (www.PlasmoDB.org), The Sanger Plasmodium Genetic Modification Project (PlasmoGEM) and PhenoPlasm (PhenoPlasm). The database has been described in 2 papers (in Trends in Parasitology and Methods in Molecular Biology).
26 September 2018
The RMgm database now contains information on the genotype and phenotype of >4490 different mutants.
27 March 2018
The RMgm database now contains information on the genotype and phenotype of >4300 different mutants.
05 September 2017
Below you will find a brief description for several updates of the RMgm database of genetically modified malaria parasites
• Update in RMgmDB of growth rate phenotypes of blood stages of PlasmoGem mutants
We have updated the growth rate phenotypes of blood stages of all mutants from the PlasmoGem resource
Changes were made to phenotypes of 234 (of the 2455) PlasmoGem mutants, and phenotypes of 117 new mutants were added.
• Linking phenotypes of RMgmDB to a new database
Phenotype data of malaria mutants (with deleted/disrupted genes) from the RMgm database have been linked to the PhenoPlasm database, a new database describing phenotypes of gene knock-out malaria parasites. See also the paper 'PhenoPlasm: a database of disruption phenotypes for malaria parasite genes.
• Phenotypes of RMgmDB mutants and PlasmoDB PlasmoDB has improved the display of phenotype data of mutants from the RMgm database (mutants with deleted/mutated genes)
• Phenotypes of new mutants in RMgmDB
A number of mutants (and their phenotypes) were recently added to the RMgm database, for example novel mutants for functional analyses of the micronemal sporozoite proteins P52 and P36 (PMID: 28506360), mutants lacking proteins involved in sporozoite motility (PMID: 28525314), mutants expressing mutated forms of thrombospondin-related protein 1 (PMID: 28115054), mutants from a knockout screen of all ApiAP2 genes (PMID: 28081440), mutants for functional analyses of proteins involved in carbon metabolism (PMID: 28027318), mutants lacking proteins that are putative targets for CDPK4 phosphorylation (PMID: 28481199).
The RMgm database now contains information on the genotype and phenotype of 4265 different mutants
02 August 2017
We have updated in the RMgm database the growth rate phenotypes of blood stages of all mutants from the PlasmoGem resource
Changes were made to phenotypes of 234 out of the 2455 PlasmoGem mutants and phenotypes of 117 new mutants were added.
21 June 2017
Phenotype data of malaria mutants (with deleted/disrupted genes) from the RMgm database has been linked to the PhenoPlasm database, a database of disruption phenotypes for malaria genes.
See also the paper'PhenoPlasm: a database of disruption phenotypes for malaria parasite genes
30 June 2017
Improved display in PlasmoDB of phenotype data of malaria mutants (with deleted/mutated genes) from the RMgm database
07 November 2016
Information on several mutants that were recently added to the RMgm database of genetically modified malaria parasites
1) A number of transgenic P. berghei mutants expressing fluorescently-tagged members of variant exported proteins of different multigene families (PIR, Fam-a and Fam-b). Analysis of these mutants showed unexpectedly that these proteins are not only expressed in blood stages and exported into the red blood cell but are also expressed in liver stages where they are exported to the parasitophorous vacuole. Evidence is presented that members of one large family (Fam-a ) transfer phosphatidylcholine. This is the first demonstration of a biological function of any exported variant protein family of rodent malaria parasites. The analyses of these mutants have been published in PloS Pathogens.
2) A large number of mutants (> 60) expressing GFP-tagged putative exported proteins GFP from a study aiming at identification of P. yoelii proteins involved in remodelling the host red blood cell. identified a significant number of conserved remodelling proteins across Plasmodium that likely represent key conserved functions in the parasite and provides new insights into parasite evolution and biology. The analyses of these mutants have been published in Scientific Reports.
3) A large number of mutants in which genes encoding transporters were targeted for gene-deletion or gene-tagging. For example 35 genes encoding (orphan) membrane transport proteins and multiple genes encoding ABC-transporters. Several ABC transporters are dispensable for blood stage growth and were shown to play a role in mosquito stages or having an essential function during liver stage development. The analyses of these mutants have been published in Mol Microbiol, Cell Microbiol and Nat Commun.
4) Mutants lacking expression of proteins involved in de novo heme synthetic pathway providing more insight into the non- essential function of this pathway during blood stage development and its role for full development of sporozoites and liver stages. The analyses of these mutants have been published in Infection and Immunity and in Mol Biochem Parasitol. See also the study on parasite heme synthesis in Plos Pathogens.
01 November 2016
We have recently added asexual blood-stage growth phenotypes for more than 2000 P. berghei mutants to the RMgm database.
The data were generated by the Malaria Programme at the Wellcome Trust Sanger Institute and reflect data in their PlasmoGem website.
These mutants were generated by transfection with a large pools of barcoded gene knockout vectors form the PlasmoGEM resource. Growth rate phenotypes were obtained by counting barcodes on a next generation sequencer daily between days 4 and 8 post transfection. It is in the nature of the screen that genotypes of individual mutants were not validated. The project website http://plasmogem.sanger.ac.uk/ has further information on methods and vector designs, provides tools for data visualisation and analysis, and allows researchers to request vectors to recreate mutants, confirm individual phenotypes and conduct more in-depth analyses.
Phenotypes were defined as follows:
• Essential: growth rate not significantly different from 0.1.
• Dispensable: growth rate not significantly different from 1 (corresponding to wild type).
• Slow: growth rate significantly above 0.1 but below 1 (p < 0.05).
• Fast: growth rate significantly above 1 (p<0.05).
In the RMgm database all genes/mutants classified as essential are classified as ‘genetic modification not successful’.
See the following link as an example: http://www.pberghei.eu/index.php?rmgm=1584
In total 1138 genes were classified as being essential using this mass screening approach.
The information of all mutants will soon be linked to the gene pages of GeneDB and PlasmoDB
The RMgm database now contains information on phenotypes of >4000 mutants
Recently a large number of mutants have been added in which proteins have tagged with GFP in a study aiming at identification of P. yoelii proteins involved in remodelling the host red blood cell.
20 May 2016
The database contains at this moment information on more than 1390 different genetically modified mutants of rodent malaria parasites.
Recently a large number of mutants have been added in which genes encoding transporters were targeted for gene-deletion or gene tagging (for example multiple genes encoding ABC-transporters and 35 genes encoding (orphan) membrane transport proteins)
05 August 2015
Information on several mutants that were recently added to the RMgm database of genetically modified malaria parasites (and information on updating P. berghei gene IDs)
1. Two P. berghei mutants (so called 'transgenic acceptor lines') that are used to create parasites in which parasite proteins can be specifically and rapidly degraded. This method employs an auxin-inducible degron (AID) and it requires the use of the acceptor lines that express the OS-TIR1 protein. This system can then be used to conditionally regulate Plasmodium protein expression.
The conditional degradation of proteins requires tagging a Plasmodium genes of interest (GOI) in the Os-TIR1-expressing mutants with the aid sequence. Subsequently, these parasites expressing both Os-TIR1 and a AID-tagged GOI need to be treated with auxin for degradation of the AID-tagged protein.
These transgenic lines have been used to analyse the function of Calcineurin, a calcium-regulated phosphatase, in multiple life cycle stages by specific degradation of this protein. See the mutants RMgm-1306 and RMgm-1307 where Calcineurin has been tagged with the aid-sequence (published in Cell, Host & Microbe)
2. Ten transgenic P. berghei mutants expressing P. falciparum vaccine candidate (sporozoites/liver-stage) antigens. These antigens are expressed under the control of the P. berghei sporozoite/liver-stage specific promoter UIS4. These mutants have then been used in immunisation/challenge studies in mice to determine P. falciparum vaccine efficacy induced by different vaccine-candidates in vivo (published in Sci Rep)
3. A P. berghei mutant which completely lacks expression of MSP7/MSP7-related proteins. This mutant has been generated by complete removal of the locus that contains 3 MSP7(-related) genes; these genes are located in a single locus on chromosome 13 in the P. berghei genome. These parasites are viable throughout the complete life cycle (published in Mol Biochem Parasitol).
4. A large number of disrupted or tagged gene mutants (mutants 1279-1301); these genes have been identified as targets of a member of the AP2-family of transcription factors; i.e. AP2-O (published in PloS Pathogens)
New GeneDB gene models/IDs for P. berghei
In RMgmDB we have now updated P. berghei gene models to the new systematic gene-IDs as provided by GeneDB in July 2015. These new gene models have been established after additional (PacBio) re-sequencing data and re-annotation of of the P. berghei genome. Though most of the core genes did not change, it was necessary to add a “0” to the gene-IDs to show differences between the versions. This has been performed by the Wellcome Trust Sanger Institute and is currently not published yet.
For RMgmDB this means that while the database can still be searched using old gene models, only the new gene-IDs will be displayed
07 May 2015
1. We have added to the RMgm database a large number of ‘kinase mutants’ (46) that were confirmed in the population of parasites, which had been transfected using a pool of PlasmoGEM vectors (vectors and mutants generated at the Sanger Institute and published in Cell Host & Microbe).
Although the kinase genes have been targeted using ‘disruption’ vectors, the mutants (or unsuccessful attempts) have NOT been classified in the database as having a ‘disrupted’ genotype (or an unsuccessful disruption). As you may know we distinguish five genotypes in RMgmDB: ‘gene disrupted’, ‘gene mutated’, ‘gene tagged’, ‘gene transgene’ and ‘gene other’. As we also did with PiggyBac insertion mutants, we have classified these mutants as ‘gene other’, since the mutant parasites have not been cloned (or analysed in extensive detail) and cannot be obtained for further analysis.
Two examples of these mutants are shown in the links below:
The parasite was generated by the genetic modification (serine/threonine protein kinase RIO1, putative) http://www.pberghei.eu/index.php?rmgm=1217
The gene/parasite could not be changed/generated by the genetic modification (protein kinase PK4) http://www.pberghei.eu/index.php?rmgm=1192
2. We have added a number of mutants (20) lacking enzymes involved in the digestion of haemoglobin in red blood cells. These P. berghei mutants were generated in a forward genetic screen targeting genes encoding these enzymes. These mutants have recently been published in Journal of Experimental Medicine. This study revealed that, unexpectedly, malaria parasites can develop without digesting haemoglobin but are restricted to young red blood cells (reticulocytes) for their development and become insensitive to the action of chloroquine.
14 November 2014 - Greatly improved genome assemblies of three rodent malaria parasites have been published.
Recently greatly improved genome assemblies of three rodent malaria parasites (RMP) have been published, together with genome wide RNAseq data of multiple RMP life cycle stages. These analyses resulted in full-length gene models for more than 98% of predicted RMP protein-coding genes. Approximately 60% of these genes have functional annotation, which is comparable to the percentage of functionally annotated genes in the P. falciparum 3D7 reference genome. A high percentage (~90%) of the predicted RMP proteins have orthologs in primate malaria species. This high level of orthology and gene conservation between RMP and primate malaria genomes further supports the use of RMP as experimental models to characterize Plasmodium gene function.
This work has been recently published in a paper entitled ‘A comprehensive evaluation of rodent malaria parasite genomes and gene expression’ by Otto et al in BMC Biology; the consortium that contributed to this study consisted of teams from the Wellcome Trust Sanger Institute (UK), RUMC and LUMC (The Netherlands), NIMR, Univ. of Glasgow and Univ. of Oxford (UK).
By sequencing additional isolates/lines of P. berghei, P. yoelii and P. chabaudi (including the subspecies P. c. adami) genotypic diversity within different RMP species has been determined in this paper. In addition, full-length chromosomal annotation permitted a comprehensive (re)classification of all subtelomeric multigene families including the largest RMP gene-family, the `Plasmodium interspersed repeat genes’ (PIR family). Both in this paper and on the ‘P. berghei SharePoint site’ it is possible to find detailed information on the origin and characteristics of different P. berghei isolates
This paper contains 17 ‘Additional Files’ with data arising from the different analyses. For example:
- Additional file RNAseq: RNAseq values of all genes in multiple synchronized blood stages, including gametocytes, and in ookinetes of P. berghei. The RNAseq data have been deposited at PlasmoDB.
- Additional file gene families: All gene members (P. berghei, chabaudi, yoelii) from 10 multigene families (mainly located in subtelomeric regions of RMP chromosomes). Several multigene families have been renamed (e.g. fam-a, fam-b, fam-c) and these names have also been updated in GeneDB and PlasmoDB
- Additional file pexel: All RMP proteins predicted to contain a PEXEL motif, identified using an updated HMM algorithm ExportPred v2.0 (J. A. Boddey et al., 2013; Traffic. 14, 532).
11 July 2014 - Last week 56 new ‘phosphatase’ mutants were added to the RMgm database.
The RMgm database now contains information on the genotype and phenotype of more than 1000 different genetically modified malaria parasites.
These 56 new mutants (gene-deletion mutants and tagging-mutants) have been generated in a systematic functional analysis of the entire P. berghei ‘protein phosphatome’, which encompasses all 30 predicted protein phosphatases (PPs). In this study, published in Cell Host & Microbe, 30 and 29 PPs were identified in the genomes of P. berghei and P. falciparum respectively, 28 of these are direct orthologs within 5 PP families. Gene disruption analysis of all P. berghei PPs revealed that half of the genes are likely to be essential for asexual blood stage development; whereas six are essential for sexual development/sporogony in the mosquito.
In addition, the first reported gene-deletion mutant in a rodent parasite that has been generated using the CRISPR/Cas9 genome editing system has been added to the database (published in Mbio by J. Yuan and collaborators)
The CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats and Cas9 endonuclease-mediated genome editing) system utilizes a prokaryotic RNA programmable nuclease that can introduce a double-strand break (DSB) at a specific site on a chromosome through heterologous expression of two components: Cas9 nuclease and a targeting single guide RNA (sgRNA). Target-specific DSBs introduced by the CRISPR/Cas9 system can be repaired by homologous recombination if a donor template is provided.
The CRISPR/Cas9 system has been shown to be highly efficient in other organisms for generating gene knock-in (KI), KO, or allelic replacements.
Click here for the description of the SERA1 gene-deletion mutant and an description of the CRISP/Cas9 constructs and technology used to delete the P. yoelii.
22 April 2014 - The database contains at this moment information on more than 950 different mutants
4 december 2013 1. New GeneDB gene models/IDs for P. yoelii
In RMgmDB we have now changed from using the previous designation of P. yoelii gene models (i.e. PYxx…) to the new systematic IDs as provided by GeneDB (i.e. PY17X_xx…..). This means:
- For all mutants only the new gene models/IDs will be visible in RMgmDB
- While the database can still be searched using old gene models, only new gene models will be displayed
For both rodent and human Plasmodium species there is an increasing depth and breadth of genome sequences available for different parasite isolates/lines. In GeneDB/PlasmoDB the (orthologous) genes from lines of the same species have been given different gene IDs, for example for the two laboratory lines, YM and 17X, of P. y. yoelii.
For all mutants in RMgmDB we standardly refer to ONLY the gene IDs of a SINGLE ‘reference line’ for each of the 3 different rodent malaria species:
- P. berghei ANKA (PBANKA_xx...)
- P. y. yoelii (PY17X_xx...)
- P. c. chabaudi (PCHAS_xx...)
In addition we link the rodent gene to the ID of the orthologous gene of P. falciparum 3D7 (PF3D7_xx...).
In the case of P. y. yoelii we provide ONLY the gene ID for P. y. yoelii 17X, whether the mutants were generated in 17X or in YM. If a mutant was generated in the YM line, we mention the YM gene ID but only as free text in ‘Additional remarks phenotype’ section.
In future we propose to use the same strategy for genes targeted in different P. chabaudi genomes (AJ, CB isolates of P. chabaudi chabaudi and isolates of P. chabaudi adami) and in different P. berghei genomes (NK65, K173, SP11).
2. Unpublished malaria mutants in RMgmDB
Recently we have included in RMgmDB a number of unpublished malaria mutants (gene knock-outs, gene-taggings etc). This includes mutants that have been generated in the laboratories of Dr. Taco Kooij and Dr. Inga Siden-Kiamos.
Researchers are encouraged to submit information to RMgmDB on mutants that are unpublished and for which no (immediate) plans exist to publish their generation/analysis. For example data on Plasmodium mutants that were generated but did not exhibit a clear phenotype as well as information on unsuccessful attempts to disrupt a parasite gene. This information (while sometimes difficult to publish) is of significant value and can also prevent an unnecessary duplication of effort (and consequently an unnecessary use of laboratory animals). The existence of null-mutants without a distinct phenotype might provide information about the functional redundancy of the target gene. Similarly, the lack of an observable phenotype might be the result of assays that are currently inadequate or, as yet, too insensitive to reveal a phenotypic effect of the genetic modification. Further (collaborative) analysis of such mutants in improved phenotype assays might reveal novel aspects of gene function. We provide a simple (excel) template for submitting to RMgmDB the information on unpublished mutants and negative trials to disrupt or mutate genes.
20 August 2013 - New information on genetically modified/transgenic parasites in the RMgmDB database (now contains information on >860 mutants)
A number of P. berghei gene-deletion or gene-tagging mutants of genes that encode (putative) palmitoyltransferases (s-acyl transferases), recently described in a paper published in Traffic (Frénal et al. 2013), have been added to the RMgm database.
See link: DHHC genes (18 mutants)
A number of P. berghei gene-deletion and gene-tagging mutants of genes that encode proteins that are part of the PTEX export machinery have been added; see the recent paper published in Mol Microbiol. (Matthews et al. 2013).
See link: PTEX genes (11 mutants)
Linking between RMgmDB and PlasmoDB and GeneDB
In PlasmoDB (www.plasmodb.org) and GeneDB (www.genedb.org) there are direct links from appropriate gene pages to the relevant genetically modified rodent malaria parasites in RMgmDB.
‘Batch searching’ RMgmDB for mutants using multiple gene ID’s.
It is possible to search and retrieve collated information related to multiple Plasmodium genes at once, which not only speeds up searches but also facilitates the cross-linking and mining of data from multiple sources.
The output for searches using multiple gene ID’s will include: Gene ID (rodent and P. falciparum); RMgm ID, the type of modification (disruption, mutation, tagging etc); if the modification was (un)successful; life cycle stages with a phenotype and if it is different or not-different compared to wild type parasites. This data will be displayed in a table on the website or it can be downloaded as an Excel/spreadsheet compatible format (i.e. as a CSV file).
Linking between RMgmDB and PlasmoGEM
In the RMgm database the information on DNA constructs used to produce mutants made using PlasmoGEM vectors has been linked directly to the detailed information available at the SANGER PlasmoGEM database (http://plasmogem.sanger.ac.uk/).
31 May 2013 - The database contains at this moment information on more thany 800 different mutants
31 december 2012 - The database contains at this moment information on nearly 750 different mutants
26 september 2012 - We have introduced improved search possibilities on RMgmDB and it is now possible to ‘batch search’ using multiple gene models. Consequently, it is now possible to search and retrieve collated information related to multiple genes at once, which not only speeds up searches but also facilitates the cross-linking and mining of data from multiple sources.
The output for searches with multiple gene ID’s will include:
Gene ID (rodent and P. falciparum); RMgm ID, the type of modification (disruption, mutation, tagging etc); if the modification was (un)successful; life cycle stages with a phenotype and if it is different or not-different compared to wild type parasites.
This data will be displayed in a table on the website or it can be downloaded as an Excel/spreadsheet compatible format (i.e. as a CSV file).
For example, searching with multiple gene ID’s will allow the user to:
- Rapidly identify genes that have been targeted for disruption but did not result in selection of mutants, indicating an essential role of these genes in blood stage development
- Identify genes that result in phenotypes in different life cycle stages, such as blood stages, gametocytes, oocysts, liver stages etc
- identify genes that have been tagged (for example with fluorescent markers)
- identify gene for which no information based on mutant phenotypes currently exists
30 July 2012 - 2. We have incorporated a module that permits linking between genotype information in RMgmDB with information on DNA-constructs available in the SANGER PlasmoGEM database (http://plasmogem.sanger.ac.uk/). Information on DNA constructs of all mutants generated using PlasmoGEM vectors will be linked, in the future, directly to the information on the PlasmoGEM database.
27 July 2012 - We have now changed from using the previous P. falciparum gene models (i.e. PFxxx.. MALxxx.., etc) to the new systematic id’s as provided by GeneDB; i.e.PF3D7_xxx..). This means:
- For all mutants only the new gene models will be visible
- The database can still be searched using old gene models, however only new gene models will be displayed
1 July 2012 - The database contains at this moment information on more than 660 different mutants
2 February 2012 - Recently a number of new Plasmodium mutants have been published in the RMgmDB database
- P. berghei mutants lacking expression of a protein, GEST, a protein indentified as being involved in infectivity of both gametocytes and sporozoites. Localization studies of GFP-tagged GEST provides evidence that this proteins is secreted by gametocytes and by salivary gland sporozoites (Talman et al., 2011 Mol Microbiol).
- P. berghei mutants generated for the ’conditional knock-down’ of AMA1 and RON4 in the sporozoite stage. Analysis of these mutants provide evidence for independent roles of AMA1 and RON4 during hepatocyte invasion
To silence the AMA1 and RON4 expression, the FLP/FRT site-specific recombination system was used to specifically delete the 3’ regulatory sequences of ama1 and ron4 in sporozoites.(Giovannini et al., 2011, Cell Host Microbe).
- RMgmDB contains several P. berghei mutants expressing tagged RON2 and RON4 proteins and also reports unsuccessful attempts to disrupt ama1, ron2 and ron4 in blood stages, indicating their essential nature for asexual blood stage proliferation
- P. berghei mutants expressing P. vivax proteinsIn these mutants the dhfr-ts gene of P. berghei is replaced with (pyrimethamine-resistant) dhfr-ts genes of P. vivax (Somsak et al., 2011 Malar J)
- P. berghei mutants lacking expression MSP7 and/or plasmepsin4, which show (virulence) attenuation during blood stage proliferation
- RMgmDB contains information on three independent P. berghei mutants lacking MSP7. All show a slightly slower blood stage proliferation, most probably due to a more severe restriction in reticulocyte invasion compared to wild type parasites. Rodent malaria parasites have three MSP7-related genes, all adjacently located on the same chromosome. The exact orthology with the 6 MSP7-related genes of P. falciparum, (MSP7 (PF13_0197) and the five MSP7-related proteins (MSRP1-5), is not clear.
- P. berghei mutants lacking expression of SMAC, a protein exported into the red blood cell; this protein is shown to be involved in CD36-mediated sequestration of P. berghei schizont (Fonager et al., 2011 J Exp Med)
In this study an additional 18 gene deletion mutants has been reported, as well as the unsuccessful attempt to disrupt a further 11 genes (therefore likely to be essential in proliferation of asexual blood stages). All mutants have been included in RMgmDB.
- Plasmodium kinases
RMgmDB contains >70 entries/P. berghei mutants generated for the analysis of Plasmodium protein kinases (see also Tewari et al., 2010 Cell Host & Microbe). All mutants have now been updated with information on the functional analysis of the P. falciparum orthologs by gene targeting (as published in Solyakov et al., 20011 Nat Commun).
- ETRAMPs
RMgmDB contains a number of entries/mutants for functional analysis of members of the ETRAMP family of both P. yoelii and P. berghei. ETRAMPs belong to a conserved Plasmodium family of proteins. These small (frequently less than 200 aa) integral membrane proteins contain a signal peptide plus a transmembrane domain and are frequently located within the parasitophorous vacuolar membrane (PVM). The ETRAMP/SEP proteins of P. yoelii and P. berghei (8-11 genes) show homology to members of the ETRAMP family of proteins of P. falciparum (14 genes) but the orthologous relationship of the different members is not completely resolved. Functional analysis of the different P. yoelii/berghei ETRAMPs by gene-tagging and gene-deletion indicate some are essential while others are not essential for asexual proliferation and demonstrate expression in both blood stages and/or liver stages, PVM location and export into the red blood cell cytoplasm (in punctuate, vesicle-like structures).(Mackellar et al., 2011. Cell Microbiol; Currà et al., 2011. Traffic).
- P. berghei parasites lines that are ’deleter lines’ for conditional mutagenesis.
These parasite lines can be used ‘to silence genes’ in the sporozoite/liver stage using the Flp/FRT site-specific recombination (SSR) system. These parasite lines have been used to silence expression of ama1, ron4, msp1 and pkg in sporozoites for the study of their role during sporozoite/liver stage development(Lacroix et al., 2011 Nat Protoc)
- ‘Promoter-swap’ mutants in which expression of a gene is specifically down regulated in gametocytes/ookinetes while expression is maintained in asexual blood stages
In the promoter-swap approach the promoter of a gene is replaced by an 'asexual blood stage-specific' promoter that is silent in gametocytes/ookinetes. 'Promoter swapping' is a powerful approach for functional analysis of proteins in gametocytes (and in subsequent stages), when the protein is normally essential during asexual blood-stage development. This approach has been used to study the function of the histone chaperone FACT in gametocytes and myosin A in ookinetes (Siden-Kiamos et al., Cell Microbiol 2011; Laurentino et al., Cell Microbiol. 2011)
- New reference P. berghei (ANKA) and P. yoelii (17XNL) ‘mother lines’ for the rapid introduction of transgenes free of a drug-selectable marker. The use of these lines (in combination with the GIMO transfection method), which greatly simplifies and speeds up the generation of mutants expressing heterologous proteins, free of drug-resistance genes, and is far faster and requires fewer laboratory animals than existing protocols (Lin et al., 2011. Plos One)
A P. yoelii 17XNL reporter line, PyGFP-luccon, which is marker-free and expresses a GFP-luciferase fusion protein under control of the constitutive eef1α promoter. Similar P. berghei reporter lines have been used to visualize and quantify host parasite interactions in vivo, analysis of drug-susceptibility, and in vivo quantification of liver stage development (Lin et al., 2011. Plos One)
29 September 2011: A new version of genome of the P. falciparum 3D7 strain is now available on GeneDB, and in this new version all genes have been assigned new gene identifiers (gene ID’s; starting with PF3D7_xxxxxxx)
For users of RMgmDB this means:
• RMgmDB can now be searched using the new PF3D7_ID’s, however only the previous P. falciparum gene identifiers will be displayed.
• We do not intend to change from the ‘old’ P. falciparum ID’s to the new systematic ID’s as displayed in GeneDB until PlasmoDB have also adopted the new ID’s
• Links in GeneDB and PlasmoDB (i.e. phenotype information) to both existing and new mutants in RMgmDB are NOT affected
Currently, RMgmDB contains information on nearly 600 different parasite mutants
6 June 2011- Recently a number of new malaria mutant parasites have been published in RMgmDB that show phenotypes with potential relevance for drug/vaccine development. P. berghei mutants expressing mutated circumsporozoite proteins
- P. berghei mutants expressing circumsporozoite proteins in which the natural H-2Kd restricted epitope is replaced with an H-2Kb restricted epitope. Phenotype analyses of these mutants indicate that the main targets of protective CD8+ T cells are parasite proteins exported to the hepatocyte cytosol. (RMgm-613)
- P. berghei mutants expressing circumsporozoite proteins with mutated ‘region I’. Phenotype analyses of these mutants establish the link between CSP cleavage and hepatocyte invasion and demonstrates that efficient cleavage requires region I (RMgm-608). P. berghei mutants expressing P. falciparum proteins
- P. berghei mutants expressing the Chloroquine Resistance Transporter (CRT) of the human parasite P. falciparum (RMgm-610, RMgm-611) - P. berghei mutants expressing the Hexose Transporter 1 (HT1) protein of the human parasite P. falciparum. These mutants have been used to test the sensitivity PfHT1 in vivo to an inhibitor of HT1, the glucose analog C3361 (RMgm-604) P. berghei mutants lacking expression of potential drug target proteins
P. berghei mutants that lack expression of ‘antioxidant enzymes’ (thioredoxin reducatase and glutathione reductase) demonstrating that these enzymes are not essential for blood stage development.(RMgm-586, RMgm-587, RMgm-403)
P. berghei mutants with disrupted genes that show a developmental defect in sporozoite development and transition into liver stages
- P. berghei mutants lacking genes encoding RNA binding proteins (Pumilio; Puf). Phenotype analyses indicate that the Puf2 protein is a key regulator of the transformation of sporozoites into liver stages (RMgm-514, RMgm-516, RMgm-616, RMgm-617).
- P. berghei mutants lacking genes encoding members of the Cysteine Repeat Modular Protein family (CRMPs). Phenotype analyses show a role in sporozoite egress from oocysts and during liver stage development (RMgm-584, RMgm-585)
24 March 2011 - The database contains at this moment information on 540 different mutants
23 March 2011 - The RMgm database now also contains ~70 new entries created by piggyBac-mediated mutations/insertions into P. berghei genes (see Fonager, J et al., BMC Genomics (2011) 12:155). Using the search term ‘piggyBac’ in RMgmBD results in a list of ~70 genes which contain piggyBac inserts. The piggyBac inserts were identified using a TAIL PCR approach in DNA collected from blood stage parasites. For all mutated genes the site of integration is indicated as ~20 nucleotides of gene sequence on either site of the TTAA insertion site. Only piggyBac insertions have been included that were found in the open reading frame, 5’UTR (0-500bp from the start codon) or within introns of genes. These piggyBac insertions are expected to affect/disrupt expression of the gene. The presence of an insert may therefore provide indirect evidence that the gene is NOT essential for blood stage development. For this reason we have included this information in RMgmDB. However, neither expression of the target gene nor the phenotype of the mutant parasite containing the insert has been analyzed.
22 October 2010 - The database contains at this moment information on more than 440 different mutants
22 October 2010 - Unsuccessful attempts to knock-out P. berghei genes based on the old gene models
IMPORTANT NOTE:
The RMgmDB contains information on a large number of (un)published and unsuccessful attempts to knock-out Plasmodium genes. These unsuccessful attempts strongly indicate that these genes may have an essential role during blood stag development. The new P. berghei annotation, in GeneDB, has revealed that for several unsuccessful attempts to knock-out genes the primers for the 5’ and 3’ targeting regions are located in NOT ONE BUT IN TWO neighboring/adjacent genes. Therefore the integration of the DNA-construct would target TWO genes. The failure to disrupt/knock-out the target gene might, therefore, be due to the unintentional disruption of an additional gene. In RMgmDB we have indicated this new information on the unsuccessful attempts where it is now apparent that the targeting construct will disrupt two adjacent genes.
22 October 2010 - RMgmDB database now also contains 64 new entries/mutants generated for the analysis of Plasmodium protein kinases, details of this study have recently been published in Cell Host & Microbe (2010, 8:377-87).
19 July 2010 - Apaper has been published in Trends in Parasitology which describes the (use of the) RMgm-database. Special emphasis is on standardization of mutant generation and describing mutant phenotypes.
26 March 2010 - In GeneDB there are direct links from appropriate gene pages to genetically modified rodent malaria parasites in the RMgm database.
5 March 2010 - We have now changed from using the previous P. berghei gene models (i.e. PBXXXXX.XX.X) to the new systematic id’s as provided by GeneDB; i.e. PBANKA_xxxxxx). This means:
- For all mutants only the new gene models will be visible
- The database can still be searched using old gene models, however only new gene models will be displayed
- The P. berghei gene models are now linked to GeneDB gene pages (and not to PlasmoDB, as it still uses the old gene models).
- Existing links in PlasmoDB out to the RMgm database are not affected. Information on new mutant entries will be linked to PlasmoDB once they have adopted the new gene models.
29 December 2009 - The database contains at this moment information on more than 300 different mutants.
23 July 2009 - In PlasmoDB there are direct links from appropriate gene pages to genetically modified rodent malaria parasites in the RMgm database.
16 June 2009 - The database contains at this moment information on more than 260 different mutants.
1 April 2009 - The database contains at this moment information on more than 200 different mutants.
28 January 2009 - The database contains information on approximately 150 different mutants.
6 November 2008 - This database is in its preliminary stages; we are still technically improving the database. At this moment we have information on less then 100 parasite mutant lines.
5 October 2008 - The new database, RMgmDB, online!
The RMgm-database contains information information on genetically modified malaria parasites generated by labs worldwide. It contains data of three rodent malaria parasites: Plasmodium berghei, P. yoelii and P. chabaudi. The aim of this database is to provide the research community access to information on the generation (e.g. disruption, tagging, mutation, transgene expression) and phenotype of mutant malaria parasites. The database also provides information on (un)published mutants without a clear phenotype and negative attempts to disrupt genes. The RMgmDB database has been described in several papers. Trends in Parasitology, Methods in Molecular Biology, Methods in Malaria Research
The information in the RMgm database is shared and linked to the database of `EuPathDB Bioinformatics Resource Center for biodefense and Emerging / Re-emerging Infectious Diseases` (www.PlasmoDB.org), the database./website of the Sanger Plasmodium Genetic Modification Project (PlasmoGEM) and the PhenoPlasm database (PhenoPlasm).
The RMgm database is maintained by Chris Janse (Leiden University Medical Center, LUMC, Leiden, The Netherlands). For questions, suggestions and remarks please contact Dr. Chris J. Janse.